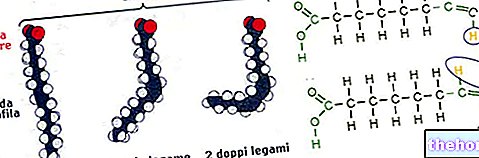
Sinteza masnih kiselina započinje iz acetil koenzima A i približno odgovara obrnutom putu njihove razgradnje; u sintezi masnih kiselina početnom acetil koenzimu A dodaje se niz bikarbonatnih fragmenata.
Sinteza masnih kiselina potpuno je citoplazmatska (tj. Enzimi koji kataliziraju ovu sintezu nalaze se u citoplazmi). Acetil koenzim A koji se koristi u citoplazmi za sintezu masnih kiselina ima mitohondrijsko podrijetlo: mali dio transportira se kroz karnitin, djelovanjem dva enzima acil transferaze (jednog citoplazmatskog i jednog mitohondrijskog) i enzima translokaze. Dio acetila koenzim A mitohondrijskog podrijetla dobiva se specijaliziranim putem: citratna liza (naziv potječe od prvog enzima ovog puta).
Acetil koenzim A prisutan u mitohondrijima potječe od glikolize nakon djelovanja piruvat dehidrogenaze; Acetil koenzim A podliježe djelovanju enzima citrat sintaze: ovaj enzim katalizira stvaranje citrata reakcijom acetil koenzima A s oksaloacetatom. Ako Krebsov ciklus može podmiriti energetske potrebe, počinje citrat (količina nepotrebna u krebsov ciklus) mogu napustiti mitohondrije i doći do citoplazme, gdje ih enzim citratna liza, trošeći energiju, pretvara natrag u acetil koenzim A i oksaloacetat. Na ovaj način moguće je imati acetil koenzim A dostupan u citoplazmi; nastali oksaloacetat mora se, međutim, vratiti u mitohondrije kako bi ponovno bio dostupan za enzim citrat sintazu.
Oksaloacetat se tada djelovanjem enzima pretvara u malat malat dehidrogenaza citoplazmatski (troši se citoplazmatski NADH): malat je propusni metabolit i može ponovno ući u mitohondrije gdje se pod djelovanjem enzima mitohondrijske malat dehidrogenaze pretvara u oksaloacetat (dobiva se i NADH); citoplazmatski pacijent može, alternativno, podvrgnuti djelovanju jabučnog enzima, koji provodi dekarboksilaciju i dehidrogenaciju, da se pretvori u piruvat. Jabučni enzim djeluje na NADP + (sličan je nikotinamid -adenindinukleotidu, ali, za razliku od toga, ima fosfornu skupinu na drugoj hidroksilnoj skupini na jednoj od dvije ribozne jedinice) pa se u prijelazu iz malata u piruvat proizvodi NADPH ( koji se koristi u biosintezi) Piruvat tada ulazi u mitohondrije gdje se djelovanjem piruvat karboksilaze ili u acetil koenzim A preko piruvat dehidrogenaze pretvara u oksaloacetat.
Pogledajmo primjer: osam molekula acetil koenzima A potrebno je za sintezu palmitinske kiseline (lanac sa šesnaest atoma ugljika), ali se samo jedna od njih koristi kao takva: sedam molekula acetil koenzima A "enzimom se pretvara u malonil koenzim A" acetil koenzim A karboksilaza (ovaj enzim koristi molekulu CO2 i ima biotin kao kofaktor).
Enzim acetil koenzim A karboksilaza može postojati u gotovo neaktivnom dispergiranom obliku i aktivnom agregatnom obliku (oko dvadeset jedinica); prijelaz iz dispergiranog u agregatni oblik događa se kada u citoplazmi postoji "visoka koncentracija citrata: citrat je pozitivni modulator enzima acetil koenzima A karboksilaze.
Enzim acetil koenzima A karboksilaze ima i druge pozitivne (inzulin) i negativne (glukagon, adrenalin i acil koenzim A) modulatore.
Analizirat ćemo sintezu masnih kiselina u bakteriji escherichia coli u kojoj se ta sinteza događa djelovanjem sedam različitih proteina; u eukariotskim stanicama mehanizam kojim se odvija sinteza masnih kiselina sličan je bakterijskom, ali se u eukariota sedam enzima odgovornih za sintezu grupiraju u dva multienzimska kompleksa A i B.
U bakterijama sedam različitih gena kodira:
- ACP (protein acil -nosač);
- ACP-acetil transacetilaza;
- ACP.malonil transacetilaza;
- β-keto-acil-ACP sintaza (kondenzacijski enzim);
- β-keto-acil-ACP reduktaza;
- D-β-hidroksi-acil dehidrataza;
- enoil-ACP redigirano.
Kod eukariota dva gena kodiraju za:
Podjedinica A
ACP;
Kondenzacijski enzim
β-keto-acil-ACP reduktaza.
Podjedinica B
ACP-acetil transacetilaza;
ACP-malonil transacetilaza;
D-β-hidroksi-acil dehidrataza;
enoil-ACP redigirano.
Sedam proteina Escherichia coli raspoređeno je tako da postoji središnji (ACP), a ostalih šest oko njega.
U njegovo enzimsko djelovanje uključene su dvije sulfhidrilne skupine: jedna koja pripada cisteinu i jedna koja pripada dugom kraku fosfopanteina; ACP se veže za supstrat koji se preko kraka fosfopanteina dovodi u kontakt s drugim enzimima koji su na taj način sposobni provesti svoje enzimsko djelovanje.
Acetil koenzim A (pomoću ACP acetil transacilaze) veže se na ACP-enzim (točnije na sumpor cisteina koji tvori cisteil-derivat) i oslobađa se koenzim A; tada intervenira ACP-malonil transakcilaza koja katalizira napad malonila na fosfopanteteinu (također se u ovom procesu oslobađa koenzim A koji je u početku bio vezan za malonil).
Sljedeći korak uključuje β-keto-acil ACP sintazu koja je kondenzacijski enzim: omogućuje fuziju između dva kostura; malonil se lako dekarboksilira i nastaje karbonil acetilnog derivata cisteina: cistein se oslobađa i nastaje derivat β-keto (acetil acetil) fosfopantetina.
Nakon toga intervenira β-keto-acil-ACP reduktaza koja reducira karbonil dalje u ACP-enzim (hidroksid stvara NADPH koji se reducira u NADP +).
Sada, 3-hidroksi-acil ACP dehidrataza djeluje (dolazi do dehidracije) što dovodi do stvaranja nezasićenog sustava (alkena).
Sljedeći postupak uključuje enoil-ACP-reduktazu (provodi hidrogenaciju: nastaje alkan, a NADPH se reducira u NADP +).
Posljednja faza uključuje pretvaranje acil produkta dobivenog u prvom ciklusu u spoj sposoban za započinjanje drugog ciklusa: enzim transakcilaze prenosi acil na cistein, ostavljajući slobodno mjesto pantetina koje će sada biti spremno vezati još jedan malonil.
U β-oksidaciji, molekula FAD-a koristi se za dobivanje nezasićenog α-β metabolita trans enoil koenzima A dehidriranjem; u sintezi masnih kiselina, umjesto toga, molekula NADPH se koristi za izazivanje suprotne reakcije.
Obično se sintetiziraju masne kiseline sa šesnaest ugljikovih atoma, ali se mogu proizvesti i masne kiseline s osamnaest, dvadeset ili dvadeset dva atoma ugljika; masne kiseline se zatim esterificiraju u tvorbu triglicerida s aktiviranim glicerolom (tj. glicerol 3-fosfatom). Potonji se može dobiti iz dihidroksi aceton fosfata djelovanjem enzima glicerol fosfat dehidrogenaza ili iz glicerola putem enzima glicerol kinaza.
Sintetizirane masne kiseline moraju se poslati u masno tkivo; transportiraju se u krvotok u obliku triglicerida ili, djelomično kao takvi, uz uporabu transportnog proteina koji je albumin.
.jpg)