UDRUŽENJE (ILI "POVEZIVANJE" ILI "VEZA")
Do sada smo liječili Mendelovo dihibridno (ili polihibridno) križanje pretpostavljajući da se različiti alelni parovi zapravo nalaze na različitim homolognim parovima kromosoma. No, broj kromosomskih parova, iako se razlikuje od vrste do vrste, varira u uskim granicama (nekoliko vrsta dosegnu oko stotinu kromosoma), dok se broj gena može izbrojiti u desetke tisuća.
Da su likovi koje je Mendel odabrao za svoje pokuse odvojili sve neovisno (bez da su stoga zbunjivali izračune u raspodjeli fenotipova u F2 polihidridnog križa) bila je sretna prilika. Ako su dva para alela pronađena na susjednim lokusima, zakon koji bi uslijedio nazvali bi se zakonom udruživanja.
Znajući da vrlo mnogo likova ima svoje mjesto na jednom kromosomu i da su oni kromosomski parovi koji se neovisno odvajaju u mejozi, može se dobro razumjeti koliko se često događa da dva para znakova, ako su bili povezani na kromosom u roditeljskom položaju podjednako i u gameti pa stoga i u organizmu u koji će donijeti vlastiti genetski materijal.
Tako vidimo da "udruživanje predstavlja" iznimku, daleko od toga da je rijetka, od neovisnosti navedenog u trećem Mendelovom zakonu.
ZAMJENA ILI "PREKRŠANJE" I PREPORUKA
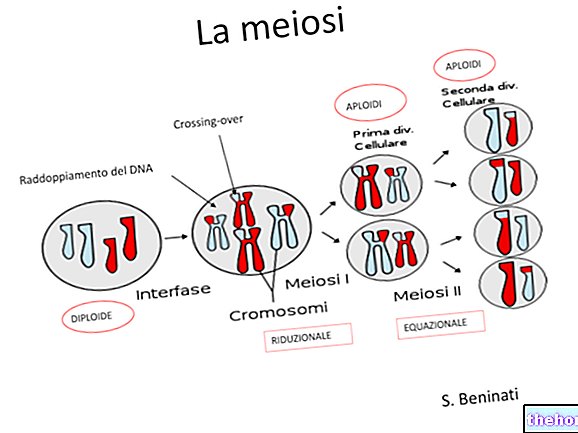
Govoreći o mejozi, naznačili smo da postoje dva različita trenutka miješanja genetskog materijala: jedan je onaj segregacije kromosoma u gametama, a to je onaj koji je primijetio Mendel.
Drugi trenutak, koji zapravo prethodi, je onaj u kojem četiri kromatide svakog para homolognih kromosoma međusobno izmjenjuju identične osobine. Nakon te razmjene dva će čimbenika koja su povezana na istom kromosomu umjesto toga biti neovisna u gametama. Vjerojatnost da se razmjena odvija razmjerna je, u prvom približavanju, duljini kromosoma, a u dužim kromosomima može biti čak i više od razmjene.
Pojava se može otkriti citološki, promatranjem dovoljnog broja mejoza pod mikroskopom.
Stopa ponovnog podudaranja je stopa po kojoj se bilo koja dva znaka koja su bila povezana u roditeljskoj generaciji međusobno rekombiniraju u F2.
Ako su dva lokusa apsolutno susjedna, vjerojatnost da će ih hiasma razdvojiti bit će praktički nula. Stopa rekombinacije bit će: n ° rekombinanata. Ako su dva lokusa na dva različita kromosoma, stopa rekombinacije bit će 0,5 (jednaka vjerojatnost da se dva znaka koja su spojena u P generaciji nasumično nađu zajedno u F2). Stopa rekombinacije stoga može varirati između 0,0 i 0,5. Za male udaljenosti na kromosomu udaljenost i brzina rekombinacije izravno su proporcionalni. Za veće udaljenosti postoji mogućnost da se dvije razmjene odvijaju između dva lokusa. Sada će se pokazati jasnim da su dva čimbenika odvojena s dvije razmjene ponovno povezana. u ovom je trenutku jasno da se gubi proporcionalnost između udaljenosti lokusa i vjerojatnosti rekombinacije.
Pronađeni lokusi povezani na istom kromosomu predstavljaju 'asocijacijske skupine'. Vrlo udaljeni lokusi mogu imati takvu vjerojatnost odvajanja razmjenom da se ponašaju kao neovisni, ali svaki od njih bit će povezan, s nižom stopom rekombinacije, s međulokusima.
Kad su poznate stope rekombinacije između mnogih parova gena unutar pridružene skupine, može se pristupiti izgradnji 'genetskih karata'. Imajući na umu da je udaljenost između dva gena (a i b) izražena brzinom rekombinacije i da udaljenost a od trećeg gena c može biti zbroj ili razlika s obzirom na udaljenost od b, moguće je rekonstruirati kartu uzajamnih udaljenosti, koja će biti genetska karta unutar te skupine udruživanja, odnosno tog kromosoma.
Sada moramo općenito razmotriti neke koncepte koji ograničavaju fenotipsku manifestaciju genotipskih obilježja.
Prije svega govorit ćemo o pojmovima penetrantnosti i ekspresivnosti, a zatim ćemo posebnu pozornost posvetiti fenomenima regulacije djelovanja gena.
PENETRANCE
Penetracija gena predstavlja njegovu sposobnost da se manifestira u fenotipu. Prodornost se statistički mjeri brojeći učestalost fenotipova koji pokazuju taj karakter od 100 genotipova koji ga sadrže. Svojstvo s penetrantom 0,7 je svojstvo koje se fenotipski javlja u 70% njegove genotipske učestalosti.
EKSPRESIVNOST
Ekspresivnost je kvantitativna procjena stupnja fenotipske manifestacije.
UREDBA RODNOG DJELOVANJA
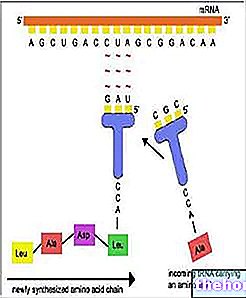
Stanice proizvode sve svoje enzime i proteine istom brzinom i u isto vrijeme. Na primjer, stanice Escherichia coli mogu se opskrbiti energijom i atomima ugljika iz disaharida laktoze jer ih mogu razgraditi na glukozu i galaktozu zahvaljujući enzimu beta-galaktozidazi. U normalnoj E. coli koja može imati laktozu, postoji približno 3 000 molekula beta-galaktozidaze, što je jednako 3% proteina te stanice; u nedostatku laktoze bit će samo jedna molekula beta-galaktozidaze po bakterijskoj stanici. Galaktozidaza će se sintetizirati iz novih molekula mRNA kad se može koristiti. Sojevi mutanata E. coli bogati enzimom poznati su čak i kad nema laktoze: ti su mutanti u nepovoljnom položaju u usporedbi s normalnim stanicama jer su prisiljeni na nepotrebnu potrošnju energije i materijala za proizvodnju enzima koji će ostati bez supstrata. Tvari koje uzrokuju povećanje količine enzima, kao što je slučaj s laktozom, zvat će se induktori, dok će se za enzime reći da su inducibilne. Druge tvari potiču, također one na specifičan način, proizvodnju određenih enzima. Također u E. coli, na primjer, sposobne izgraditi sve svoje aminokiseline, koje imaju ugljik i amonij (NH3), prisutnost u mediju za kulturu određene aminokiseline (na primjer histidin) blokira proizvodnju svih enzima povezanih s biosinteza same aminokiseline: za ove će se enzime reći da su represivni.U bakterijskim stanicama molekule mRNA ruše se nedugo nakon njihova formiranja i zato kontrolirati proizvodnju mRNA znači kontrolirati enzimsku sintezu u isto vrijeme vrijeme.
OPERON
Kako bi objasnili kako bakterijska stanica može kontrolirati vlastitu proizvodnju enzima, Jacob i Monod formulirali su hipotezu o operonu; operon tvori nekoliko gena koji su funkcionalno povezani i poravnati bez diskontinuiteta duž proteza DNK.Operon se sastoji od tri različite vrste gena: promotora, gdje počinje stvaranje mRNA, operatora, gdje se vrši kontrola; jedan ili više strukturnih gena koji kodiraju enzime ili druge proteine. U sustavu beta-galaktozidaze, operon uključuje, osim onoga za beta-galaktozidazu, i dva druga strukturna gena koji kodiraju druge enzime koji sudjeluju u metabolizmu laktoze. Ti su geni susjedni jedan drugome i prepisuju se jedan na drugi jedan za drugim duž iste spiralne DNK u jednoj molekuli mRNA. Tako proizvedene molekule mRNA aktivne su vrlo kratko, nakon čega ih uništavaju specifični enzimi.
Aktivnost operona s druge strane kontrolira drugi gen, regulator, koji također može biti udaljen od operona: ovaj regulator kodira protein, nazvan represor, za koji se čini da se veže za DNA u genu operatora. između promotora i strukturnih gena zapravo blokira proizvodnju mRNA.
Potiskivač se pak kontrolira, a kontrola se vrši pomoću "signalne" tvari. U slučaju inducibilnih enzima ova tvar je "induktor. Induktor" se veže na molekulu represora mijenjajući svoj oblik tako da se više ne može prilagoditi DNK: u ovom slučaju, budući da ne postoji represor između promotora i strukturnih gena , represor može tvoriti molekule mRNA i od njih proteinske molekule. S iscrpljenjem opskrbe induktorom, regulator će ponovno uspostaviti kontrolu, što će zaustaviti proizvodnju nove mRNA, dakle novih proteina. U sustavu beta-galaktozidaze induktor je laktoza ili tvar vrlo slična ovoj. Derivat: oni pridružit će se represoru deaktivirajući ga tako da omogući biosintezu enzima. U slučaju represivnih enzima, tvar koja djeluje kao "signal" djeluje kao corepressor: represor je aktivan samo ako se kombinira s corepressom. U histidinskom sustavu, koji uključuje desetak različitih enzima, ta je aminokiselina u kombinaciji sa svojom tRNA, jezgrom potiskivačem.
ALOSTERIČKI INTERAKCIJE
Alosteričke interakcije, uključujući inaktivaciju enzima promjenom njegovog oblika, pružaju drugačiji način regulacije metaboličke aktivnosti stanice. Alosteričke interakcije omogućuju precizniju kontrolu od induktor -represorskog sustava operona, ali ne postižu koristan rezultat isključivanja biosinteze određene tvari iz prve faze - proizvodnje mRNA.
SUSTAVI UPRAVLJANJA U EUKARIJU
Postoje neke činjenice koje navode na zaključak da sustav regulacije sličan operonu djeluje i prevladava među biljkama i životinjama. Kromosomi ovih organizama uvelike se razlikuju od onih E. coli i drugih prokariota. Kontrola gena u ovim stanice su vrlo različite.Mehanizam mitoze je takav da svaka stanica određene biljke ili životinje posjeduje sve informacije
genetika prisutna u oplođenom jajašcu. Stoga će većina gena u bilo kojoj specijaliziranoj stanici ostati neučinkovita tijekom cijelog života stanice. DNA u tim stanicama uvijek je povezana s proteinima. Dakle, moguće je da represija gena u eukariota zahtijeva upravo ovu povezanost između DNK i proteina.